

















1. Introduction
Photosynthesis research has a long history, dating back to at least the 17th century. In a sense, a special biological reaction called photosynthesis involves a physical process called light energy conversion and a chemical process called redox, and its elaborate mechanism is of interest to many researchers, not only in the biological field. I pulled it. In addition to this, in the 21st century, the promotion of research on photosynthesis has been strongly desired due to some external factors. In other words, in order to break away from the dependence of energy on oil and nuclear power, promotion of the use of natural energy such as solar energy is an urgent issue, and photosynthesis research as a model system for photoenergy conversion becomes important and biomass. Photosynthesis research is also attracting attention as a measure to improve production efficiency. Looking at the macro aspect, the mechanism of photosynthesis and carbon cycle, which plays a role in absorbing carbon dioxide in the environment in the global ecosystem, is clarified on a global scale while global environmental problems are being highlighted. It is becoming desirable to do so. In addition to research on the mechanism of photosynthesis, it can be said that research on the use of photosynthesis is becoming necessary. In this paper, based on these changes in the situation, I would like to introduce the progress of photosynthesis research focusing on some points and think about the future of the research.
2. Photosynthesis research so far
The basic composition of photosynthesis, which uses the energy of light to break down water and generate oxygen to fix carbon dioxide to organic matter, was already revealed at the end of the 18th century. Regarding the specific mechanism, the pathway of carbon assimilation that fixes carbon dioxide to organic matter was clarified in the 1950s, and the main reaction components of the basic mechanism of light energy conversion were identified in the 1960s. .. Since it became possible to handle membrane proteins by biochemical methods with surfactants in the 1970s, the properties of protein complexes responsible for photosynthesis have been elucidated, and since the 1980s, molecular biology has progressed. The method has made it possible to unify the mechanism of photosynthesis beyond biological species. The results of photosynthetic reaction center crystallization and X-ray crystallography, which won the Nobel Prize in Chemistry in 1988, became the starting line for research methods that reduce functions to structures for discussion. To date, the three-dimensional structure of major proteins including the photosystem reaction center complex responsible for photosynthesis has been clarified, and the detailed structure of the manganese cluster responsible for water decomposition of the photosystem II reaction center has not been known until the end. The structure was finally revealed this year with a resolution of 1.9 Å. It seems that the players in the game of photosynthesis are now almost clear.
Under such circumstances, what direction is the current photosynthesis research aiming for? One direction would be the dynamic aspect of how individual players are bargaining with each other in the game of photosynthesis. The time aspect of responding to the external environment is important for this, and many of these remain unclear. Among them, the response to light that directly affects photosynthesis, that is, the response to the optical environment, has been studied as a central theme. In addition, the global photosynthesis research mentioned above has also become possible from simple estimation to discussion based on data due to the development of remote sensing by satellite. However, I will revise the manuscript on these topics, and first, I would like to introduce the structure determination of the photosynthetic reaction center as a recent research example of photosynthetic research.
3. Structure and function of photosynthetic reaction center
(1) Two photosystems

In oxygen-evolving photosynthesis performed by land plants, algae, and cyanobacteria, there are two types of photosystem reaction center complexes responsible for photoenergy conversion, which are called photosystem I and photosystem II, respectively. These photochemical systems consist of 20 to 30 protein subunits and about 100 molecules of complementary molecules such as photosynthetic dyes bound to them, because they are hydrophobic supermolecular complexes that penetrate the photosynthetic membrane system. Although structural analysis was delayed, the work of crystallizing the reaction center of photosynthetic bacteria, which won the Nobel Prize, and structural analysis became a breakthrough, and research is progressing rapidly in recent years.
Photosystem I has a relatively low oxidation-reduction potential of the electron transport chain in the complex, and plays a role in reducing ferredoxin and NADPH, which act as reducing agents in biochemical reactions in vivo. Photosystem I was crystallized in cyanobacteria in 1998, and structural analysis progressed in the early 2000s. The photosystem I of land plants has an extra antenna complex called LHCI around the reaction center complex, so its structural analysis was delayed, but the crystal structure was clarified in 2003.
On the other hand, Photosystem II is responsible for the decomposition reaction of water. In order for water to break down and release oxygen, four electrons must be extracted from two molecules of water. The difficulty of water splitting is that the reaction does not proceed unless the oxidizing power of 4 electrons is accumulated, and the only biological reaction that can proceed this reaction is the photosystem II reaction center complex of the photosynthetic system. Oxygen evolution reaction in. In the oxygen evolution of photosynthesis, the accumulation of oxidizing power for 4 electrons is carried out by manganese clusters containing 4 atoms of manganese bound to the photosystem II reaction center complex. The structure of this manganese cluster was unknown until the end, but the detailed structure was finally revealed this year as described below.
(2) Structure of oxygen evolution system
While the crystal structure of the photochemical reaction center complex has been clarified in various biological species, the resolution of the crystal structure of the manganese cluster, which is the water decomposition site of the photosystem II reaction center complex, has not been sufficient so far. The detailed structure was unknown. However, very recently, the structure of the photosystem II reaction center complex was clarified with a resolution of 1.9 Å, and the structure of the manganese cluster was determined, including the coordination of surrounding water molecules. This structure was determined for Thermosynechococcus vulcanus, a type of cyanobacteria that is a unicellular prokaryotic photosynthetic organism. You can think of it as having the same structure.

According to the reported structure, four manganese atoms, together with one calcium atom and five oxygen atoms, form a "squashed chair" structure as an Mn4CaO5 cluster (Fig. 1). Three of the manganese atoms, four of the oxygen atoms, and a total of eight calcium atoms make up the dice-shaped part, and the remaining manganese and oxygen atoms make up the backrest. Calcium atoms are located in the upper part of the chair, not on the backrest side, and coordinate two water molecules with manganese atoms in the backrest. The reason why the chair is crouched is that the distance between the oxygen atom present at the base of the backrest and the surrounding calcium atom and the two manganese atoms is longer than that between other atoms. Therefore, it is considered that this oxygen atom exists in a state closer to the hydroxide ion, which indicates that this oxygen atom may be derived from one of the substrates water. Since the calcium atom existing in the vicinity of the oxygen atom and the manganese atom on the backrest coordinate the water molecule as described above, the water molecule serving as another substrate is one of these. There is a possibility.
(3) Redox state of oxygen evolution system
Kok's "oxygen clock" model has been proposed for the process of accumulating the oxidizing power of four electrons in manganese clusters, and this is still almost accepted at this time. This assumes five S states S0 through S4 in the manganese cluster, here,

Each time one electron is extracted from the manganese cluster by light irradiation, the S state transitions from S0 to S1, S1 to S2, S2 to S3, and S3 to S4.
The S4 state has a short lifespan, releases oxygen molecules and returns to the S0 state, and is in a state in which two molecules of water, which are substrates, are coordinated.
One proton is released during the transition from S0 to S1 and one from S2 to S3, and two protons are released during the transition from S4 to S0.
When left in the dark for a long time, the S2 and S3 states transition to the S1 state in the opposite direction.
Given this behavior, we can generally explain the experimental facts that have been obtained regarding water splitting. Manganese atoms in manganese clusters are generally considered to have Ⅲ and Ⅳ valences (in the S0 state, Ⅱ valences are also possible), and oxidizing power accumulates as changes in the valences of the four manganese atoms. do. It is considered that the crystal whose structure has been analyzed is mainly in the S1 state in the process of crystallization in the dark. Structural information in other S states is essential to clarify the dynamics of the water splitting process, but unfortunately, structural analysis of reaction center complexes in other S states is technically difficult at this time.
4. Photosynthetic efficiency

(1) Light absorption efficiency
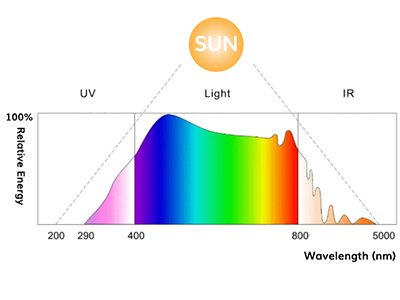
Whether aiming at artificial photosynthesis or using photosynthesis for biomass production, the premise is high photosynthesis efficiency of plants. Then, how high is the efficiency of photosynthesis in plants? In order to use light for photosynthesis, it must first be absorbed. The light absorption rate by the leaves of a plant varies depending on the type and wavelength of the plant, but can be considered to be 80% or more on average. In recent years, closed plant factories with artificial lighting have become a kind of boom, but at this time, green light that chlorophyll does not absorb is inefficient, so red LED and blue LED are used for growth light. There are examples of increasing efficiency. Certainly, looking at the absorption spectrum of chlorophyll in an organic solvent, the absorption of green light near 550 nm is about two orders of magnitude smaller than the absorption of blue light and red light at the maximum absorption limit. Looking at this, it is not unreasonable to think that plants cannot absorb green light, but in reality, the absorption rate of green light in the leaves of plants exceeds 70% on average for various plant species.
This high value is achieved by the high chlorophyll content of the leaves and the placement of cells in the leaves. When the cross section of the leaf is observed with a microscope, cylindrical cells are regularly arranged vertically on the surface side. While there is air between cells, the inside of the cell can be regarded as water as an approximation, so due to the difference in refractive index, these cells act as optical fibers and guide light to the inside. On the other hand, when approaching the back side of the leaf, amorphous cells are irregularly arranged, and as a result, light is scattered and returned to the inside of the leaf without being transmitted to the back side as it is. With such a mechanism, the optical path length of the light transmitted through the leaf is up to several times the thickness of the leaf, and even green light can absorb a considerable part of it. On the contrary, in the case of green light, the cells inside the leaf also contribute to photosynthesis, whereas in the case of green light, most of the light is absorbed by the surface cells of the leaf when the light is strong enough. Therefore, the efficiency of photosynthesis is even higher in the case of green light.
In any case, the amount of light absorption increased by irradiating the leaves with light excluding green light instead of white light is at most a few percent, and the quantum yield of photosynthesis is blue light rather than green light, as will be described later. Therefore, it is considered that the difference in photosynthetic rate is smaller than this. Therefore, the effect of light color on plant growth through the efficiency of photosynthesis is considered to be extremely small in practice. However, plants use blue light receptors that absorb light near 450 nm and red light / near-infrared light receptors that absorb light near 670 nm / 730 nm to adjust the morphology, etc. for sensing the light environment. Therefore, when a plant is grown except for specific light, it is often observed that the morphology and the like are greatly affected, and as a result, the growth is changed.
(2) Transfer efficiency of excitation energy
The light energy absorbed by the photosynthetic dye (antenna dye) is transferred to the reaction center chlorophyll by excitation energy transfer. Dozens to hundreds of dye molecules are bound to the reaction center complex, and the excitation energy is passed between these antenna dye molecules one after another. Even if the molecular species are the same, these antenna dyes have different energy ranks due to the difference in coordination with the protein, but the difference in the ranks is not large, and it is considered that energy transfer occurs with each other at room temperature. Therefore, when focusing on a specific dye, it is considered that there are many possibilities for the route of excitation energy transfer from the dye to the reaction center chlorophyll, but the mechanism of the route selection is not always clear. In 2007, it was shown that in two types of photosynthetic bacteria, at least at liquid nitrogen temperature, multiple photosynthetic dyes (bacteriochlorophyll in this case) are in an electronic resonance state. In 2010, it was shown that photosynthetic bacteria and subsequently Cryptophyceae (the photosynthetic pigment in this case is phycobilin pigment) have similar quantum states at room temperature. The phycobilin pigments of Cryptophyceae have an average center distance of about 20 Å, but it is extremely interesting that resonance is observed between such separated pigments and even at room temperature. It is considered that the excitation energy from each antenna dye is efficiently collected in the reaction center by such resonance. Quantum mechanical effects are often thought to be observed only in controlled homogeneous systems at very low temperatures, but in life phenomena such as photosynthesis, quantum mechanical effects can be seen under very natural conditions. Will be. There is a movement to call research on such phenomena as quantum biology and consider it as a model for technologies such as quantum computing.
(3) Quantum yield of photochemical reaction

When the reaction center is excited by excitation energy transfer, an electron transfer reaction (charge separation) from the reaction center chlorophyll to the primary electron acceptor occurs. The quantum yield of this initial charge separation is 98% or more, which is extremely high. In solar cells, charge recombination after charge separation contributes to the decrease in yield, but in photosynthesis, multiple receptors that receive electrons one after another from the primary electron receptor should be placed in the reaction center complex. Achieves a high quantum yield.
The electron transfer reaction of photosynthesis involves two types of photochemical reaction center complexes (photosystem I and photosystem II) and the electron transfer components that connect them. Since electrons are extracted from water in Photosystem II and the oxidized water becomes oxygen molecules, the amount of oxygen generated is often used as an index of electron transfer activity. In many cases, the quantum yield of oxygen generation is about 10 photons to generate one molecule of oxygen, and the yield is about 80% when compared with the theoretical value of generating one molecule of oxygen with 8 photons. When the spectrum is observed in detail, the yield decreases in the blue light region near 480 nm, and the yield is rather high in the green light region where the absorption itself is small. The wavelength region where the yield decreases is just near the absorption band of carotenoids. Previously, carotenoids were thought to act as "accessory pigments" as antennas that absorb light and transfer excitation energy to chlorophyll, but recent studies have shown that β-carotenoids and zeaxanthin play a major role in light capture. It has come to be thought that it is not a collection, but rather an energy dissipation and removal of active oxygen under strong light conditions. The decrease in quantum yield in the blue light region is considered to reflect this energy dissipation process.
(4) Energy yield of photosynthesis
Next, let's look at the energy yield of the entire photosynthetic reaction. The theoretical maximum yield when light energy is fixed as chemical energy of organic matter by photosynthesis is about 30%. On the other hand, when the actual amount of light energy conversion efficiency of an actual plant is actually measured as the accumulated organic matter amount (biomass), it is up to about 5% under human control even under the conditions of a laboratory where the environment is controlled. It is said that the maximum is about 1% in some agricultural land, and it is less than 0.1% under normal outdoor conditions. Considering that the energy conversion efficiency of commercially available solar cells is about 10-20%, these values seem to be extremely low, and do not match the image of photosynthesis efficiency, which is usually regarded as "high".
There are three main causes for this. The first is the issue of "manufacturing costs." In the case of solar cells, the energy conversion efficiency is measured with the pre-manufactured one installed. On the other hand, in the case of plants, not all of the profits of photosynthesis can be accumulated as photosynthetic products, and some, and in some cases, most of them must be devoted to their own growth. It also costs money to maintain life activity, that is, to simply live. That is, in the case of plants, the manufacturing cost and maintenance cost are subtracted from the efficiency of photosynthesis, which is the main cause of the above difference between 30% and 5%. To make a fair comparison, even in the case of solar cells, the manufacturing and maintenance costs, and in some cases the disposal costs, would have to be deducted from the conversion efficiency.
The second cause is changes in the natural environment. In nature, environmental factors such as light, temperature, humidity, and soil moisture content are constantly changing, and it can be said that there is almost no time when photosynthesis can actually show maximum efficiency. Plants have various photosynthetic regulatory mechanisms for environmental changes, but in reality most of these regulatory mechanisms are "reducing the efficiency of photosynthesis." Excessive light energy is dangerous to living organisms, so under adverse environmental conditions where the rate of photosynthesis is slow, it is necessary to activate a mechanism that reduces light absorption and an energy dissipation system. That is, plants must grow for many hours with the efficiency of photosynthesis actively reduced. This is the main reason for the difference between 5% and 1% above.
The third cause is the involvement of other organisms. The accumulation of organic matter in plants in the natural environment is lost due to the feeding of other organisms. This is a feeding damage for crops, but when considering the entire global ecosystem in which plants are responsible for primary production, it can be considered as an essential part of the ecosystem cycle. In order for non-photosynthetic organisms, including humans, on the earth to survive, some of the organic matter accumulated by plants through photosynthesis must be eaten. This part, along with the second part, causes the above 1% and 0.1% difference.
Considering the above, a value of 30% would be appropriate for the energy conversion efficiency without subtracting the manufacturing cost and maintenance cost. Even in that case, this value cannot be said to be particularly high as compared with solar cells and the like. A major feature of the energy conversion system of plant photosynthesis is not its high efficiency, but its self-proliferation ability and self-repairing ability. And even plants pay a reasonable cost for this self-renewal and self-repair. In the above explanation, one point to note is that in the study of photosynthesis, light that can be absorbed by chlorophyll (photosynthetically active radiation, 400-700 nm) is targeted. If you are discussing the light absorption rate and energy conversion efficiency for light in a wide wavelength range including infrared light in research on solar cells, etc., the yield of photosynthesis described here will be used. It cannot be compared directly.
5. Aiming to improve the efficiency of photosynthesis in plants
(1) Necessity of research on the utilization of plants
At present, the development of alternative means to replace the use of fossil resources has become an unavoidable problem due to multiple aspects such as the seriousness of global environmental problems, the depletion of fossil resources, and the anxiety about relying on nuclear energy for energy. At that time, it is actually necessary to develop two types of alternative means. That is, the development of energy and the development of raw materials for the organic industry. Regarding the former, for example, the use of solar cells and the like can be considered, and technically, the stage of practical application has been sufficiently entered. On the other hand, research on making organic industrial raw materials from carbon dioxide is also being actively pursued, but for this, another technical breakthrough will be required for practical use. On the other hand, the method using plant biomass is excellent in that light energy conversion by photosynthesis and fixation of carbon dioxide to organic matter are performed at the same time, and energy and organic industrial raw materials can be obtained at the same time.
A lot of research and development of alternative energy and alternative organic industrial raw materials utilizing the power of such photosynthetic organisms have already been carried out in reality. In principle, many of the projects currently being studied are completely new, and the key to practical use is whether or not the cost can be reduced to the level of fossil resources. On the other hand, in many cost calculations, the fact is that photosynthesis efficiency, including photoenergy conversion efficiency, is increased and improved. Since the growth of photosynthetic organisms depends entirely on photosynthesis, it is natural to think that improving the efficiency of photosynthesis is important for improving the productivity of plants and algae. However, in reality, there is no actual example of increasing the efficiency of photosynthesis and increasing productivity. The main reason is that photosynthesis is a system adapted to the natural world. It is hard to think of the reason why photosynthesis is inefficient and profitable. The photosynthetic efficiency of plants adapted to the habitat should have been optimized during evolution. If the photosynthesis efficiency is not apparently optimized and a modification that artificially increases the photosynthesis efficiency is possible, the modification does not occur in the process of evolution, so the modification is in terms other than photosynthesis. It is likely to cause negative changes that counteract it. This is considered to be the reason why photosynthesis was not successful in the past at a practical level to increase productivity.
(2) When will production efficiency increase?
For example, breeding to make fruits huge does not cause the above-mentioned problem of optimization of evolutionary process. Making the fruit larger than necessary is detrimental to the plant itself and is not an option during evolution. Such changes, that is, changes that are "lossy" for plants and "profitable" for humans, can be used for breeding. However, if this is the case, improvement by breeding cannot be expected for traits that are directly linked to productivity (= fitness), such as the efficiency of photosynthesis.
If so, why did crop productivity increase in the 1960s? Around this time when the population in Asia increased rapidly, many highly productive varieties, mainly rice and wheat, were developed and called the "Green Revolution." The varieties bred during the Green Revolution vary, but the biggest change in traits that is directly linked to productivity is shortness. This made it resistant to lodging and made it possible to increase the number of ears. However, short traits are difficult to select naturally in nature because they are a negative factor for competition for light. In other words, that is why there was room for improvement through breeding. In fields and rice fields, weeding has freed us from the competition for light, and the administration of nitrogen fertilizer has made it easier for plants to grow. The Green Revolution was successful because it selected mutations that bridged the gap between natural and cultivated conditions.
(3) Is it possible to increase the efficiency of photosynthesis itself?
Considering the above, we have to think that it is very difficult to breed in a way that simply increases the maximum efficiency of photosynthesis. As mentioned in the previous section, the maximum efficiency of photosynthesis is optimized by various "devices", and it seems impossible to modify this to improve efficiency. On the other hand, photosynthetic efficiency in the field is suppressed for various reasons. Plants that grow in a fluctuating natural environment develop various mechanisms of environmental response. It should be noted that the photoenvironmental response alone can be regulated at various levels such as regulation of pigment amount, regulation of pigment protein complex balance, regulation of energy distribution between complexes, and regulation of electron transfer. Most of these are responses that reduce the efficiency of photosynthesis. That is, it is considered that plants are adapted to the natural environment by optimizing the efficiency of photosynthesis under optimum conditions and combining this with a regulatory mechanism that reduces the efficiency of photosynthesis. If so, the only way to increase the efficiency of photosynthesis is to consider a combination of environmental control and breeding to create a regulatory mechanism that can be considered as a cost for adaptation to the natural environment in a controlled environment. Isn't it to cultivate the omitted plants? In reality, we have succeeded in obtaining a gene mutant that exhibits photosynthetic efficiency superior to that of the wild type under specific circumstances. I think that such a direction may be considered as applied research of photosynthesis.
6. Conclusion
Now that the reaction center chlorophyll protein complex, which is a huge chromoprotein complex involved in photosynthesis, has been crystallized and its structure has been clarified, the target of the next research will be its dynamics. As mentioned above, the maximum efficiency of plant photosynthesis is probably optimized during the evolution of long organisms, and there seems to be only room for improvement in efficiency in dynamic control systems. The history of photosynthesis research is long, and the fact that a lot of structural and functional information has been clarified in that history does not indicate the end of photosynthesis research, but provides the basis for future dynamics research. It is a thing. Furthermore, it seems likely that research on photosynthesis will be the starting point for a new field such as quantum biology. Research on photosynthesis has been carried out in collaboration with the fields of physics and chemistry, not limited to the field of biology. On the extension line, I think it is desired to promote photosynthesis research that is not limited to a narrow field and is not divided into basics and applications.